中国更新世末全新世初广谱革命的新视角:水洞沟第12地点的动物考古学研究
张乐①*,张双权①,徐欣①②, 刘德成①,王春雪①③,裴树文①, 王惠民④,高星①
① 中国科学院古脊椎动物与古人类研究所,中国科学院脊椎动物演化与人类起源重点实验室,北京 100044;
② 中国科学院大学,北京 100049;
③ 吉林大学边疆考古研究中心,长春 130012;
④ 宁夏文物考古研究所,银川 750001
* E-mail: zhangyue@ivpp.ac.cn
收稿日期:2012-05-18
接受日期:2012-11-12
中国科学院知识创新工程重要方向项目(编号: KZCX2-EW-QN110)、中国科学院战略性先导科技专项(编号:XDA05130302)、国家重点基础研究发展计划(编号:2010CB950203)和“优秀青年人才培养”专项资助
摘要:从动物考古学角度,依照成本与收益模式,根据猎物体型大小以及躲避猎捕的速度等标准将水洞沟第12地点的动物划分为高回馈率的大型动物和低回馈率的小型慢速动物以及小型快速动物三类,利用Simpson指数对三种类型动物数量分布的均衡度进行检测,发现第12地点远古人类对三类猎物的选择倾向性较低,即肉食食谱的均衡度较高与Stiner的研究结果对比显示,第12地点的均衡度变化应该不是气候改变造成,而可能是肉食广谱革命的结果。与西亚地区的 Natufian中石器文化相关数据的详细对比,则进一步证明了这一结论。
关键词:广谱革命 水洞沟第12地点 动物考古学 更新世末全新世初 中石器时代
20世纪60年代末,Binford[1]和Flannery[2]提出了广谱革命的概念,他们认为在新石器时代之前出现一个阶段,这一时期狩猎采集人群的食谱变得宽泛,增加了新要素,因而提高了更新世之末冷暖多变的环境对人类的负载能力概念的提出主要依据12~8ka BP欧洲中高纬度地区远古人类狩猎和食物加工方式的巨大变化,出现了对植物种子研磨以及对小动物的利用等行为。此后, Bar-Yosef 和Meadow[3]根据储藏设施、保存的植物遗存等,论证了西亚新旧石器过渡期间人们对植物遗存利用的变化。Wringht[4]根据西南亚遗址出土的磨制工具,认为植物尤其是坚果和大种子植物是这一时期生计革命的必要组成部分。崔天兴[5]和Unger-Hamilton[6]则认为 Natufian中石器时代的石刀是用来收割农作物的。上述证据主要从植物性要素增加的角度证明了更新世末全新世初欧洲及西亚、西南亚地区的广谱革命;Stiner和Munro[7]则另辟蹊径,对地中海Levant 南部遗址出土的动物骨骼进行了研究,为这一地区广谱革命的发生提供了系统的动物考古学证据,并为世界其他地区广谱革命事件的确认提供了独特的研究视角。
水洞沟第12地点的年代在10ka BP左右,也出土了类似的间接证据——石磨盘、石磨棒和石杵[8],暗示了这一时期存在对植物种子进行捣碎和研磨的行为,而可镶嵌石叶成为复合工具的刻槽骨器则也可能是被当作石刀收割植物的用具。磨制石器和复合工具在年代较早的水洞沟第1,2,7和8地点均未发现,从器物组合上分析,水洞沟第12地点的古人类可能已经拓宽植物性食谱,因为利用研磨等方式对食物进行加工要耗费更多的时间和精力,在食物较为充裕的条件下古人类不会选择这样的资源[9]。虽然在距离第12地点不远、年代较早的水洞沟第2地点也发现了古人类利用植物种子的证据,但是后者的器物组合中未见石磨盘、石磨棒和石杵,仅见残留着禾本科植物种子淀粉粒的打制石器[9],这意味着水洞沟第12地点的远古人类可能对植物种子进行了更有效的加工,从加工器具上讲,已经起码具有了向原始农业过渡的要素。那么从动物资源的利用上看,水洞沟第 12地点又具有怎样的特性呢?对该遗址出土动物骨骼进行系统研究和对比,将对探讨中国晚更新世末全新世初是否发生了广谱革命提供重要的线索。
1 材料
水洞沟遗址位于宁夏回族自治区银川市东南28km处, 行政区划属灵武市临河乡,平均海拔约1200m,地处银川盆地东部边缘、毛乌素沙地西缘。水洞沟遗址第12地点位于边沟河下游“春旺砖厂”附近,在水洞沟第1地点以北约4km处,地理坐标为38°19′40.0″N, 106°29′49.0″E. 这一区域内共发育了6级阶地,SDG12号地点和其他遗址都埋藏于T2地层中。该地点2005年夏发现,2007年发掘,文化层中出土了大量石制品、骨制品和动物碎骨[10]。石制品包括细石核、细石叶、刮削器、局部磨光的石器、石磨盘、石磨棒和石杵等;骨制品包括制作精美的骨锥、骨针和可镶嵌石叶、有刻槽的复合骨制工具等[8]。出土动物骨骼万余件,所属动物包括:兔(Lepus sp.), 狗獾 (Meles meles),小野猫(Felis microtus),鹿(Cervidae),普氏羚羊(Procapra przewalskyi),野猪(Sus sp.),普氏野马(Equus przewalskyi),水牛(Bubalus sp.)以及鸟类、爬行类和啮齿类。
研究者根据堆积物分布和组成推测水洞沟第12地点的遗物为原地埋藏,略经流水的改造[10],痕迹观察也显示,动物骨骼表面未见流水磨蚀以及搬运过程中形成的摩擦痕迹,因此这些骨骼应该不是流水聚集的。该地点仅发现两件具有食肉动物咬痕的动物骨骼,比例为 0.1%,远远低于食肉类聚集的动物骨骼组合比例(82%~15%)[11];具有啮齿类啮咬痕迹的标本也仅出土两件,比例为0.1%,远远低于啮齿类洞穴中具有磨牙痕迹的标本比例(22%~100%)[12],据此可以排除是食肉类和啮齿类将动物骨骼带入遗址的。此外,有5.1%(92件标本)的动物骨骼表面存在人工切割痕,这一比例看似较低,但对于古人类聚集的动物群而言,在欧洲、非洲的遗址中这一数值大多低于 5%[13,14];在中国,经过系统埋藏学观察的灵井动物骨骼组合(旧石器时代中期)以及马鞍山遗址上文化层(旧石器时代晚期)和下文化层(旧石器时代中期)动物骨骼组合的切割痕迹出现比例分别为 13%[15],4.74%和13.17%[16]。因此,水洞沟第12地点动物骨骼组合看似较低的切割痕比例不能排除这一动物骨骼组合应该是人类聚集的,结合与动物骨骼共出的上万件石制品、大量骨器以及用火证据,我们认为该处应该是远古人类生活的遗迹。
2 水洞沟第 12 地点的动物种属分布
生态学设想生物觅食遵循两大原则,最高效地摄食和保证成功繁殖。在这个前提下,一种食物是否被获取并不取决于它在环境中的丰富性,而是以觅食效率的指标即回馈率来衡量。回馈率一般以觅食者在单位时间内获得的卡路里计,觅食者总是追求回馈率最大化的目标,即用最短的时间获得最多的食物,这就是最佳觅食理论(optimal foraging theory)的原理[17]。譬如,大型动物较难猎捕,但是其营养物质产出量较高,相对来讲就具有较高的回馈率;而那些体型较小、逃逸速度较快的动物(如兔和鸟类等)则既难猎捕又难处理,而且获得的营养物质又较少,其回馈率相应的也就很低。猎食者如果能比较容易地猎取到回馈率较高的猎物,他们就会选择放弃低回馈率的猎物。而当高回馈率的猎物资源相对变少,狩猎者对猎物的关注范围就会扩大,食谱中低回馈率的猎物比例即会增加,继而造成了更多样化或者更均衡的食谱。考古研究中被复原的肉食食谱往往采用林奈的分类系统来表示,种和属是最常用的分类单元,因此,衡量上述变化看似最直接的动物考古学证据即食谱中是否出现更多的种属或者各种属的组成比例是否出现均衡性[7]。
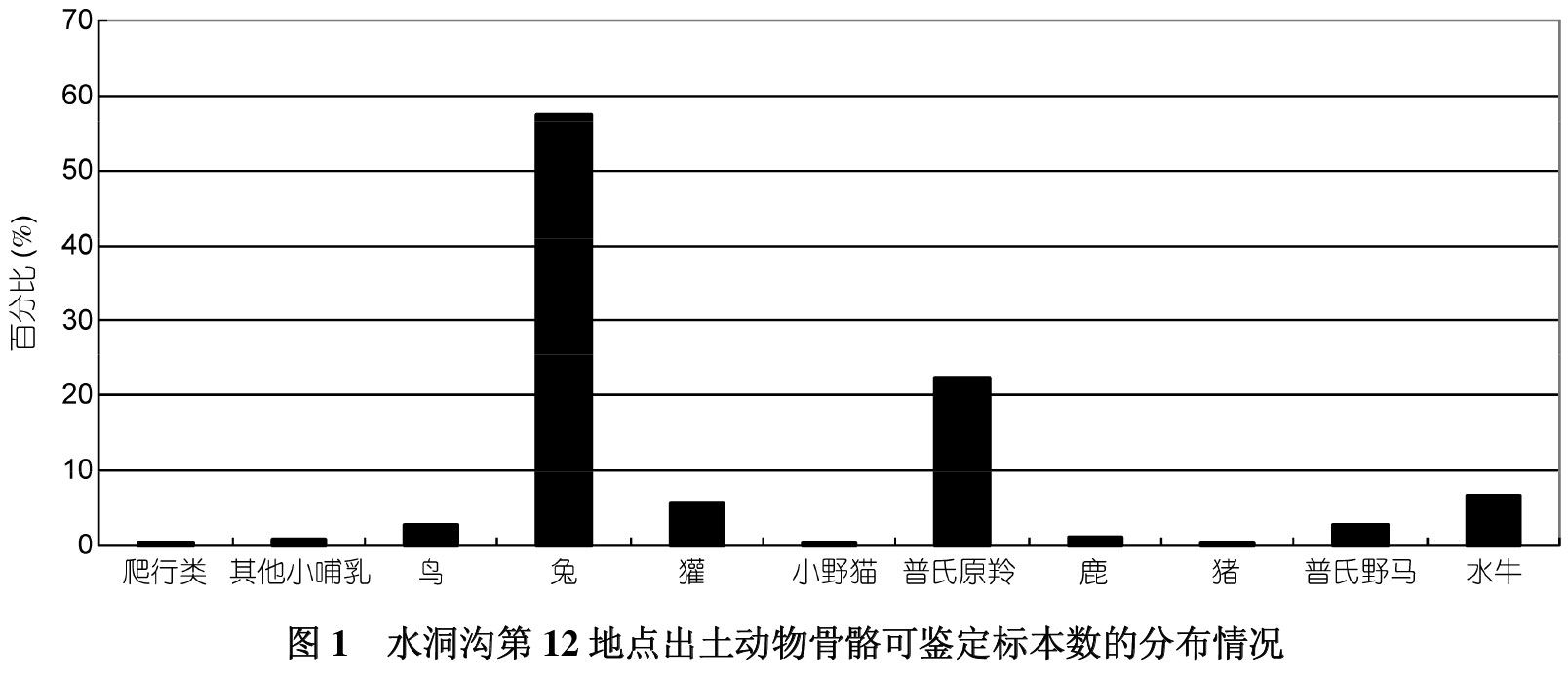
水洞沟第12地点出土动物骨骼可归为11个种属,而同一地区年代稍早的水洞沟第2地点和第7地点则均为7个种属,说明水洞沟地区稍晚阶段远古人类的肉食食谱中确实出现了新的种属,那么其分布的均衡性如何呢?第12地点可鉴定到种属与部位的动物骨骼1821件;属于兔的骨骼1045件、狗獾103件、小野猫2件、鹿19件、普氏羚羊404件、野猪5件、普氏野马52件、水牛124件、鸟类53 件、爬行类1件,其他小哺乳类13件;其中兔子的骨骼数量占优势,比例为57.39%;普氏羚羊的骨骼数量居其次,比例为22.19%;其他动物骨骼数量的比例均小于10%(图1)。这样的比例分布是否均衡呢?Simpson指数[18,19]能够对动物种属分布的均衡度进行数量化的检测。公式为1/∑(pi)2,其中p代表该动物骨骼组合中每种猎物的 NISP占NISP总量的比例,i代表这一动物骨骼组合中种属的数量。当指数值最大达到i时,种属分布最均衡;指数值最小为1,此时种属分布最不均衡,所有数值集中分布在一个种属上。种属的均衡度检测旨在揭示猎人是否会主要猎取他们偏爱的猎物,而忽略另外一些猎物,或者当时空变化后,这种偏爱是否会发生改变,那些以前从未或者偶尔被捕猎的猎物是否会成为食谱中占据一定比例的组成部分。水洞沟第12地点动物骨骼组合的Simpson指数值为3.05(分布最平均时数值为11),要低于年代稍早同属水洞沟遗址的第7地点的数值 4.85(分布最平均时数值为7),即水洞沟第12地点各种属分布的均衡度低于第7地点,如上所述,衡量肉食广谱革命看似最直接的动物考古学证据之一就是食谱中各种属的组成比例出现均衡性趋势,那么这一结果是不是说明时代处于晚更新世末全新世初的第12地点未发生肉食的广谱革命呢?
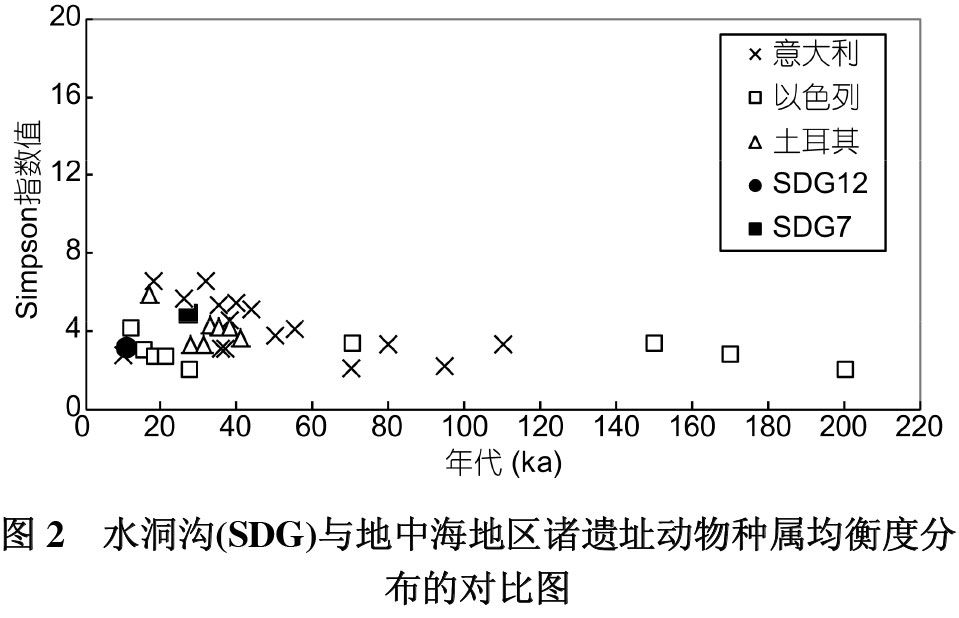
上述方法实质上是对古人类猎物选择均衡度的量化,数据对比针对的不是具体种属的多少,而是选择行为的变化。所以可以暂时抛开时空的不同以及种属的差异,将水洞沟第12地点动物骨骼组合与现有的可对比数据进行对比研究,以探讨该遗址远古人类猎食行为的特性。Stiner等[7]对地中海地区属于北部以色列(200~11ka BP)、意大利西海岸(110~9 ka BP)和土耳其中南海岸(41~17ka BP)的32个遗址动物种属分布的均衡度进行了检验,发现随着时间的变化,诸遗址动物种属分布的均衡度并无明显的规律性变化。究其原因,她认为林奈的方法虽然在生物分类上非常强大,能够为动物命名,可以指出不同物种之间的区别与联系,却不适用于人类行为的研究,因为人类捕猎追求的是觅食效率即回馈率,而并不必遵循猎物的生物学分类。根据林奈的原则,很多在分类上差异很大的动物在躲避捕猎的行为上却比较相似,其处理成本(handling cost)也类似,如野兔和鹧鸪在分类上属于不同的纲,但其体型大小和肉量相当,躲避危险的速度相似,因此其捕猎成本和处理成本基本相同。由于远古人类捕猎并不受生物学分类束缚。其变化也就很难敏感地表现在种属均衡度的变化上[7]。
所以,林奈体系下复原的食谱中出现更多的种属或者各种属分布的相对均衡并不能像想象中那么简单而直接地对肉食广谱革命进行证明[7]。水洞沟第12地点、第7地点的数据点均与同时期地中海地区某些遗址动物种属分布均衡度的数值分布几乎重合(图2),说明其种属均衡度分布与地中海地区相比并无明显差异,二者的均衡度对比也不能为该地区是否发生了肉食广谱革命做出解答。
3 水洞沟第12地点动物群新分类
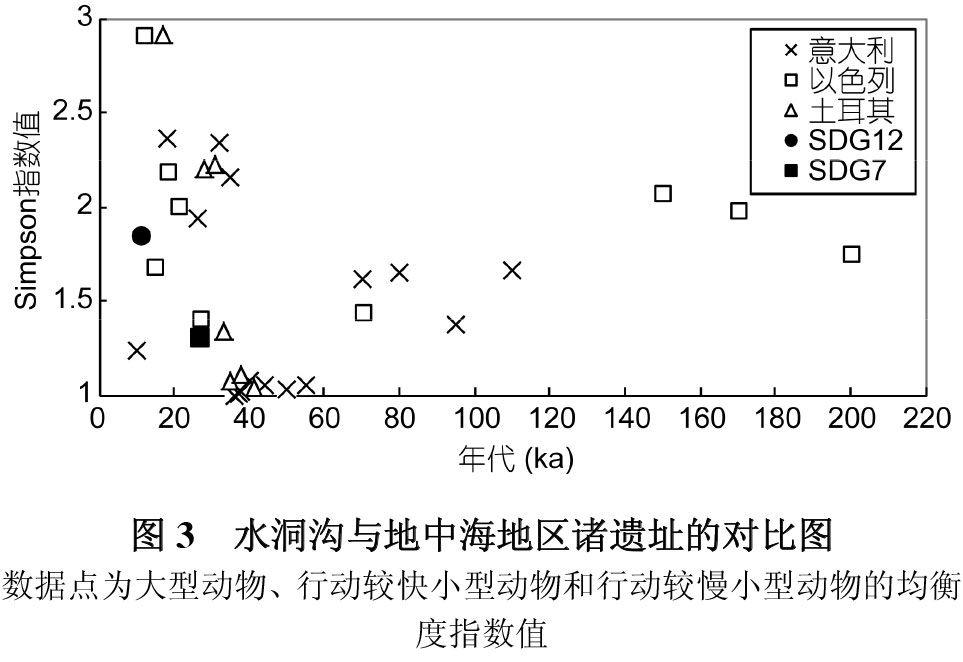
由于林奈的生物分类学方法对人类选择猎物行为及其变化的反应缺乏一定的敏感性,Stiner等[7]用成本与收益模式,根据猎物的体型大小、躲避猎捕的速度等标准将它们划分为高回馈率(大型动物)和低回馈率动物(小型动物),并进而依据生殖特点及躲避捕猎能力将小型动物进一步划分为两类:行动较快和行动较慢的动物。在新的分类系统下,上述32个遗址的Simpson指数分布具有一定的规律性(图3),在10ka左右环境暖湿时期大部分遗址三类动物的均衡度值相对较高,甚至达到最高,表明在气候环境较为适宜的情况下,远古人类对回馈率较低的猎物进行一定程度的开发[7,20],研究者认为这可能主要与此时人口激增导致资源相对减少有关[20~22]。
依据该分类, 在水洞沟第 12 地点属于高回馈率的大型有蹄类动物包括鹿、普氏羚羊、野猪、普氏野马和水牛,属于低回馈率行动较快的动物包括兔和鸟类,未发现低回馈率行动慢的龟类和贝类等,其Simpson指数值为1.84,在图上显示远远高于水洞沟第7地点的数值,说明其大型动物、行动较快小型动物和行动较慢小型动物的比例分布要比第7地点平均,即第12地点远古人类对三类猎物的选择倾向性降低,从这种意义上讲,其肉食食谱的均衡性增加。水洞沟第 12地点年代为10ka,处于更新世与全新世的交界,孢粉分析显示在其堆积时期气候暖湿[10];而在第7地点地层堆积时期,水洞沟地区植被覆盖度低,气温转冷干[23];从理论上讲二者的均衡度变化趋势与环境变化趋势相悖,即环境变暖,资源相对丰富,远古人类比较容易猎捕高回馈率的动物,均衡度应该变低,而水洞沟第12地点的情况则与之相反。因此,我们推测第12地点和第7地点三类动物分布均衡度的差异应该与气候无关,前者的均衡度指数高于后者在一定程度上可能表明在水洞沟,12地点发生了肉食广谱革命。
现阶段,由于水洞沟地区出土大量动物骨骼的地点并不多,我们尚不能在该区域扩大时空范畴的对比,从而进一步对上述结论进行证明,但与Stiner等[7]的研究结果对比显示,水洞沟第12地点、第 7地点的数据点均与同时期地中海地区某些遗址的数值分布点几乎重合(图3),说明其均衡度分布特性与趋势变化可能与地中海地区相比并无明显差异,这在一定程度上暗示水洞沟第12地点可能与地中海地区同时代的遗址一样发生了肉食广谱革命。
4 水洞沟第12地点的小型动物
地中海地区从旧石器时代中期早段开始远古人类就开始利用各种小型动物,Stiner等[7]将这些小型动物分为三类,包括回馈率相对稍高的慢速动物,如龟、贝类等,回馈率较低的快速奔跑型动物,如兔等,回馈率较低的快速飞行动物,如鸟等。通过Simpson指数计算,Stiner等[7]发现随着时间的转移,地中海的远古人类对这三种肉食资源的利用呈现越来越均衡的态势,进一步证明广谱革命在旧石器时代晚期之末开始发生。
然而,对于水洞沟第12地点来讲,按照上述分类计算的Simpson指数值很低,仅为1.1(最低值为1),因为在该遗址尚未发现回馈率相对稍高的慢速动物,如龟、贝类等。这是否为水洞沟第12地点发生了肉食广谱革命提供了反证呢?这一问题需要从环境差异的角度进行解答。水洞沟遗址地处银川盆地东部边缘、毛乌素沙地西缘,植被稀疏,雨水较少,年均温8℃,年降水量200mm,属于荒漠草原环境[23],而中国的龟鳖类则生活在多水的区域,在沿海地区其种类和数量明显高于相邻内陆地区[24];Stiner等[7]研究的32个遗址则均处于地中海地区,接近水源,有利于龟鳖的繁育,即使在旧石器时代中期大型动物资源相对丰富,该地远古人类也猎捕龟类,因为它们非常容易获得。所以,水洞沟第12地点未出土行动缓慢的龟类、贝类等动物应该是该地区缺水因而缺乏此类资源造成,而并不是捕猎范围没有拓宽。
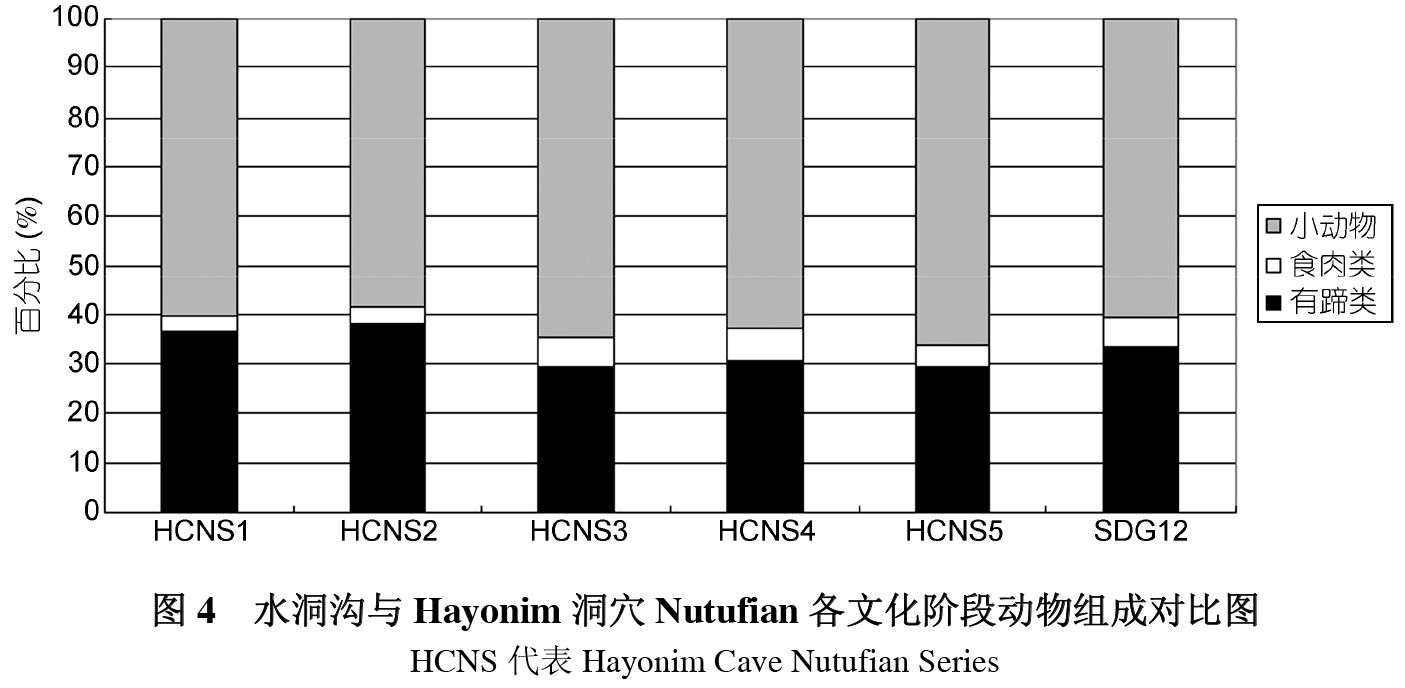
水洞沟第12地点的食谱中虽然缺乏龟类、贝类等行动缓慢小型动物,由兔和鸟类组成的小型动物所占数量和比例则不可小觑,已达到61.07%,这一比例与Levant地区的Natufian文化及其相似。Natufian文化是西亚地区最具代表性的中石器时代文化, 年代处于 12800~10200BP之间,文化组成包括细石器、磨制骨器、原始农业、装饰品、艺术和有意识的埋葬等,并被证明对资源进行了强化利用,发生了广谱革命[25]。Stiner等[7]对Hayonim洞穴Natufian文化层的动物骨骼进行了研究,将动物组成划分为有蹄类、食肉类和小动物三类,发现Natufian五个文化阶段小动物的组成比例基本都在 60%左右。这一结果与水洞沟第12地点极其相似,而与第7地点相比则大为不同,后者小动物的比例仅占13%(图4)。
研究显示人口爆炸从西南亚向欧洲延伸了多次, 遗址中鸟类骨骼数量的激增, 以及龟类骨骼的缩减是旧石器时代晚期人口扩散的最早证据[20-22]。对野兔资源的大量开发则是人口爆增在稍后阶段的标志,这在伊比利亚、东欧、北欧以及北非都有体现[26~28]。例如,在葡萄牙旧石器时代中期的人很少猎捕野兔,晚期的人只猎捕一定量的野兔,而到了中石器时代被捕猎的野兔数量则激增[27]。维基百科上记载兔子6~7个月性成熟,怀孕期只有30天,一年可产9窝,每窝可产4~12只幼兔,也就是说一只母兔一个生育季可以拥有800 只后代。所以,兔子是非常成熟和稳定的肉食资源,晚更新世末全新世初考古遗址中兔类骨骼的激增可能是远古人类应对人口暴增导致资源匮乏的结果,因而可以成为这一时期是否发生肉食广谱革命的重要证据。水洞沟第12地点兔类骨骼所占比例为52.72%,远远高于年代稍早的第7地点 (12.07%)。这也在一定程度上进一步为水洞沟第12地点发生了肉食广谱革命给予了证明。
5 结论
水洞沟第12地点大型动物、小型慢速动物以及和小型快速动物的均衡度与年代稍早的水洞沟第7地点相比较高,与同时期地中海地区诸遗址的均衡度分布特性和变化趋势相比无明显差异,而且能够提供稳定肉食资源的兔类骨骼比例激增,指示了这一地区肉食广谱革命的发生。水洞沟第12地点在文化性质上与其年代稍早的水洞沟第2地点和第7地点相比均有较大差异,在后两个遗址中未发现细石器、精致磨制骨器以及磨制石器。水洞沟第12地点石磨盘、磨棒和石杵的发现指示了一定规模的植物利用行为存在,这在一定意义上显示这一时期的远古居民可能不仅仅拓宽了肉食食谱,他们对植物性食物的利用可能也更深入和广泛。相关的残留物分析和植物遗存研究尚在进行,将由专业人员另行发表。水洞沟第12地点远古居民对肉食性和植物性资源利用的广谱革命在一定程度上揭示了这一时期该地区人类与资源的关系,环境指标显示其变化应该不是气候改变造成,而可能与世界上其他地区相似,这一阶段水洞沟地区的人口也出现暴增的现象,资源相对匮乏,因此远古人类开始较大规模地开发以前相对忽略的小型猎物资源以及较为深度地加工植物资源,发生了广谱革命。
致谢: 感谢宁夏考古研究所钟侃、马晓玲,中国科学院古脊椎动物与古人类研究所陈福友、罗志刚、张晓凌、周振宇、关莹、彭菲、李锋、仪明洁、牛东伟等在试掘、发掘及材料整理过程中给予的帮助和贡献。
参考文献:
[1]Binford L R.Post-Pleistocene adaptions. In: Binford S R, Binford L R,eds. New Perspectives in Archaeology.Chicago: Aldine Publishing Company, 1968. 313–341
[2]Flannery K V.The domestication and exploitaion of plants and animals.In: Ucko P J,Dimbleby G W,eds.The Rise and Fall of Civilizations: Modern Archaeological Approaches to Ancient Cultures. New York: Aldine Publishing Company, 1969
[3]Bar-Yosef O,Meadow R H. The origins of agriculture in the Near East. In: Price T D, Gebauer A B,eds.Last Hunters,First Farmers:New Perspectives on the Prehistoric Transition to Agriculture.Santa Fe:School of American Research Press,1995.39–94
[4]Wringht K L.Ground-stone tools and hunter-gatherer subistence in Southwest Asia:Implication for the transition to farming.Amer Ant,1994,59:238–263
[5]崔天兴.“广谱革命”及其研究新进展.华夏考古,2011:119–152
[6]Unger-Hamilton R. The Epi-Palaeolithic Southern Levant and the origins of cultivation. Curr Anthropol,1989,30:88–103
[7]Stiner M C,Munro N D.Approaches to prehistoric diet breadth,demography,and prey ranking systems in time and space. J Archaeol Method Theory,2002,9:181–214
[8]高星,王惠民,刘德成,等.水洞沟第12地点古人类用火研究.人类学学报,2009, 28:29–336
[9]关莹,高星,李峰,等. MIS 3晚期阶段的现代人行为与“广谱革命”:来自水洞沟遗址的证据.科学通报,2012,57:65–72
[10]刘德成,陈福友,张晓凌,等.水洞沟12号地点的古环境研究.人类学学报,2008,27:295–303
[11]Blumenschine R J.An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages.J Archaeol Sci,1988,15: 483–502
[12]Brain C K.The Hunters or the Hunted? An Introduction to African Cave Taphonomy.Chicago:University of Chicago Press, 1981
[13]Gaudzinski S.On bovid assemblages and their consequences for the knowledge of subsistence patterns in the Middle Palaeolithic.Proc Prehist Soc,1996,62:19–39
[14]Cain C.Human activity suggested by the taphonomy of 60 ka and 50 ka faunal remains from Sibudu cave. Southern Afr Hum,2006,18:241–260
[15]张双权,李占扬,张乐,等.河南灵井许昌人遗址动物骨骼表面人工改造痕迹.人类学学报,2011:313–326
[16]张乐.马鞍山遗址古人类行为的动物考古学研究.博士学位论文.北京:中国科学院古脊椎动物与古人类研究所,2008
[17]潘艳,陈淳.农业起源与“广谱革命”理论的变迁.东南文化,2011: 26–34
[18]Simpson E H.Measurement of diversity.Nature,1949,163:688
[19]Levins R.Evolution in Changing Environments:Some Theoretical Explorations.Princeton:Princeton University Press,1968
[20]Stiner M C,Munro N D,Surovell T A,et al.Paleolithic population growth pulses evidenced by small animal exploitation.Science, 1999, 283:190–194
[21]Stiner M C.Paleolithic population growth-response. Science,1999,284:1468–1468
[22]Stiner M C,Munro N D,Surovell T A.Paleolithic population growth pulses evidenced by small animal exploitation. Am J Phys Anthropol,2000: 292–293
[23]刘德成,王旭龙,高星,等.水洞沟遗址地层划分与年代测定新进展.科学通报,2009,54:2879–2885
[24]周婷. 中国龟鳖动物的分布. 四川动物,2006,25:272–276
[25]Bar-Yosef O.The Natufian culture in the Levant,threshold to the origins of agriculture.Evol Anthropol,1998,6:159–177
[26]Kuhn S L,Stiner M C.The antiquity of hunter-gatherers.In: Panter-Brick C,Layton R H,Rowley-Conwy P A,eds.Another Day, Another Camp:An Interdisciplinary View of Hunter-gatherers. Cambridge:Cambridge University Press, 2001.99–142
[27]Hockett B S,Bicho N F.The rabbits of Picareiro cave:Small mammal hunting during the late upper palaeolithic in the Portuguese Estremadura.J Archaeol Sci,2000,27:715–723
[28]Jochim M. A Hunter-Gatherer Landscape:Southwest Germany in the Late Paleolithic and Mesolithic.New York:Plenum, 1998
本文出自:《中国科学:地球科学》 2013年第4期,628-633页。
